Rev. Fac. Agron. (LUZ). 1998, 15: 319-329
Obtainment of antiserum against bean southern mosaic virus through a simple methodology
Recibido el 27-10-1997lAceptado el
20-04-1998
1. Parte del Trabajo Especial de Grado presentado por el primer autor para optar al
Título de Ingeniero Agrónomo en la Facultad de Agronomía de la Universidad Central de
Venezuela.
2. Dirigir correspondencia a: Prof. Mario José Garrido, Universidad Central de Venezuela,
Facultad de Agronomía, Instituto de Botánica Agrícola, Sección de Fitopatología,
Apartado 4579, Maracay 2101-A, Venezuela.
Resumen
Palabras claves: Virus, purificación, antisuero, caraota, Phaseolus vulgaris.
Abstract
Key words: Virus, purification, antiserum, bean, Phaseolus vulgaris.
Introducción
La caraota (Phaseolus vulgaris L.) es la leguminosa de mayor importancia destinada al consumo directo de la población en Venezuela. Este cultivo es afectado por numerosos patógenos, entre los cuales los virus revisten especial interés debido a las pérdidas que ocasionan y porque el control químico de los mismos es generalmente ineficaz (9).
Más de cuarenta virus han sido reportados infectando a la caraota, de los cuales el virus del mosaico sureńo de la caraota (bean southern mosaic virus: BSMV) es uno de los más diseminados mundialmente. Esta virosis puede causar pérdidas considerables en el rendimiento (más del 50%), debido a la reducción del número y peso de la semilla producida por las plantas de caraota infectadas (9). En Venezuela es poca la información existente sobre este virus y su incidencia se ha incrementado notablemente en los últimos ańos, particularmente en el estado Aragua (8, 12).
Una vía rápida y segura de identificar los virus de plantas es a través de pruebas serológicas. Sin embargo, para este tipo de prueba se requieren antisueros específicos, y para producirlos se necesitan insumos y equipos de costo elevado (15). Esta situación ha obligado, en la mayoría de los casos, a que en nuestro país deban importarse antisueros a un precio alto, lo cual dificulta los trabajos de diagnóstico y epidemiológicos, no sólo en caraota, sino también en otros cultivos. Lo anterior se debe, fundamentalmente, a la escasez de laboratorios de virología dotados de los equipos y materiales requeridos para este tipo de trabajo.
En Venezuela se han realizado algunas investigaciones dirigidas a la obtención de antisueros mediante esquemas sencillos de purificación con resultados muy halagadores (1, 11, 14).
Estos protocolos, desarrollados para algunos virus de interés en el país, podrían ser utilizados en laboratorios con limitaciones de equipos, no sólo en Venezuela, sino también en otros países con limitaciones económicas o en vías de desarrollo.
En atención a lo antes expuesto, y en vista de la importancia que tiene la caraota en nuestro país, se consideró de interés realizar esta investigación, cuyo objetivo fue obtener antisuero contra el BSMV mediante la implementación de una metodología sencilla de purificación.
Materiales y métodos
Fuente de virus y mantenimiento de plantas. Como fuente del virus fueron utilizadas plantas de caraota cv Tacarigua infectadas mecánicamente con un aislado identificado previamente como BSMV raza B (BSMV-B) (12).
Las semillas fueron sembradas en una mezcla estéril de suelo y arena, en proporción 3:1 (v/v), respectivamente, contenida en materos plásticos de 1 L de capacidad (4-5 semillas/matero). Las plantas se inocularon mecánicamente (10) a los 7-8 días de edad y se mantuvieron en un umbráculo libre de insectos, a 26-28 şC y 65-70% HR, donde fueron regadas diariamente, cada 15 días fertilizadas con una solución de una fórmula completa (16-16-08; 2-3 g/L) y como medida preventiva adicional se efectuaron aplicaciones semanales (1/2 dosis comercial) de insecticida (Anthio), acaricida (Omite) y fungicida (Polyram-Combi), permitiendo mantenerlas en un estado óptimo de crecimiento.
Purificación parcial del virus. Para purificar parcialmente al BSMV-B se siguió el procedimiento descrito en la figura 1. Este protocolo está basado en la metodología de Christie et al. (5) para concentrar partículas virales, con algunas modificaciones de los autores. La siguiente descripción agrega algunas particularidades del procedimiento: las plantas fueron cosechadas a los 19-21 días después de la inoculación mecánica (10), momento para el cual evidenciaban síntomas característicos de la infección viral; sus hojas más jóvenes fueron cortadas finamente y congeladas a -15 şC hasta su utilización (2-3 días después de la cosecha).
Las hojas eran trituradas en una licuadora en presencia del buffer, en una relación 1:2 (p/v), durante 5 min.
En el transcurso de la purificación se utilizó una centrifuga Sorvall RCL-B con un rotor SS-34. Todo el proceso se realizó a una temperatura de 2-4 şC y no se hicieron ajustes de pH.
Pruebas de infectividad. Fueron realizadas con las siguientes fracciones de la purificación: a) sobrenadante descartado después de la precipitación con polietilenglicol; b) último sedimento descartado; y c) suspensión parcialmente purificada (figura 1). Cada una de estas fracciones fue inoculada mecánicamente en plantas jóvenes (7-8 días de edad) de Phaseolus lunatus L. cv Henderson Bush, en el cual el BSMV-B induce lesiones locales necróticas (13). También fueron inoculadas plantas de P. vulgaris cv Tacarigua, la cual manifiesta síntomas típicos de mosaico ante la infección con este virus (8).
Figura 1. Esquema de purificación parcial del BSMV-B.
Obtención del antisuero. Para obtener el antisuero contra el BSMV-B se inmunizó un conejo de la raza Nueva Zelanda de 52 días de edad. Previo a la inmunización se obtuvo el suero normal, con la finalidad de constatar que el conejo a inmunizar no presentaba anticuerpos contra el BSMV-B. Además este tipo de suero es utilizado en las pruebas serológicas como control.
a) Suero normal. Para obtener el suero normal, al conejo se le afeitó la oreja derecha, se le desinfectó con alcohol etílico al 70%, se le dilataron las venas con xilol y se le aplicó vaselina para evitar que la sangre se dispersara en el borde de la oreja. El corte se realizó en la vena marginal, próxima a la base de la oreja, permitiendo que la sangre goteara hacia tubos de vidrio estériles. Seguidamente la sangre colectada se dejó durante 3 h bajo condiciones de laboratorio. Al cabo de ese tiempo se separaron cuidadosamente los coágulos de las paredes de los tubos utilizando una varilla de vidrio estéril; los tubos fueron colocados en un refrigerador a 4 şC hasta el día siguiente (3, 15).
El suero fue separado de los coágulos y centrifugado a 3000 g durante 15 min. Por cada mililitro de suero normal se le agregó una gota de azida de sodio al 1% como preservativo y se guardó congelado a -15 şC hasta su utilización (3).
b) Inmunización del conejo y tratamiento del antisuero. A los tres días después de la extracción de la sangre para obtener el suero normal se inició el proceso de inmunización. Al conejo se le suministraron cinco inyecciones intramusculares que contenían 1 mL de suspensión viral + 1 mL de adyuvante incompleto de Freund a intervalos de seis días (3,15). La mezcla de antígeno y adyuvante se efectuó minutos antes de cada inyección.
La extracción de la sangre se realizó de la siguiente manera: a) Un sangrado preliminar para conocer si el animal había producido anticuerpos suficientes; éste se efectuó cuatro días después de la tercera inyección. Se siguió el procedimiento descrito en la obtención del suero normal. b) El sangrado propiamente dicho se efectuó a los 11 días después de la última inyección, mediante punción cardíaca (3). En ambos casos la sangre fue tratada en la forma descrita para la obtención del suero normal.
Al suero normal y al antisuero obtenido contra el BSMV-B se les determinó el título a través de pruebas serológicas de doble difusión en agar (2). Para ello se hizo reaccionar diferentes diluciones de los sueros con diferentes diluciones de los antígenos, y se determinó la máxima dilución del antisuero capaz de detectar su homólogo (15).
c) Absorción o saturación del antisuero. Para absorber o saturar el antisuero se preparó una suspensión de proteínas de planta sana, macerando hojas jóvenes de plantas de caraota (19-21 después de la siembra) en un mortero frío y estéril, sin adición de otras sustancias; el extracto era filtrado a través de gasa y centrifugado a 3.000 g por 15 min. A partir del sobrenadante fueron preparadas varias diluciones (1/2, ... 1/16) con buffer fosfato de potasio 0,001 M, 0,85% NaCl, pH 7,5; cada una de ellas fue mezclada con igual volumen de antisuero (0,2 mL) y mantenidas a 4 şC durante 12 h. Luego, las diluciones fueron centrifugadas a 3.000 g por 15 min para eliminar precipitados. Cada dilución se probó contra proteína de planta sana a través de pruebas de doble difusión en agar, con el fin de determinar la dilución más alta capaz de absorber todos los anticuerpos contra este tipo de proteínas presentes en el antisuero. Una vez absorbido el antisuero, fue mezclado con glicerol en proporción 1:1 (v/v) y almacenado a -15 şC.
Serología. Se utilizó el método de doble difusión en agar o de Ouchterlony (2). El medio de inmunodifusión consistió de 0,8% de agar purificado, 0,85% de cloruro de sodio y 0,1% de azida de sodio en agua destilada. Una vez preparado el medio, 12-15 mL eran vertidos en cada caja de Petri plástica de 100 mm x 15 mm. Los huecos realizados en el gel medían 5 mm de diámetro y estaban equidistantes entre si, separados 5 mm de un hueco central de igual medida (2).
Cuando se utilizó material vege-tal infectado como antígeno, las hojas jóvenes de plantas de caraota (19-21 días después de inoculadas) fueron maceradas en un mortero frío y estéril, sin agregar buffer, y el extracto era filtrado a través de gasa y centrifugado a 3.000 g por 15 min. El sobrenadante era utilizado en las pruebas. El testigo era obtenido de igual forma, pero a partir de plantas sanas.
Microscopía electrónica. Para corroborar la forma y tamańo de las partículas virales, así como para comprobar la concentración viral en la suspensión parcialmente purificada, se procedió de la siguiente manera: sobre una rejilla de cobre cubierta con una película muy delgada de nitrocelulosa (colodión) y carbón evaporado se colocó una gota de suspensión viral diluida y se dejó durante 1 min. Luego, se eliminó el exceso de líquido de la rejilla con un papel filtro, se coloreó con una gota de acetato de uranilo y después de 2 min se eliminó el exceso de colorante con un papel filtro. Una vez seca la rejilla, aproximadamente a los 15 min, se observó al microscopio electrónico de transmisión (5, 10).
Resultados y discusión
En plantas de caraota `Tacarigua' el virus se multiplicó eficientemente, lo cual se evidenció en las preparaciones a partir de la suspensión parcialmente purificada observadas al microscopio electrónico. Una alta concentración de partículas isométricas de 28-30 nm de diámetro, características del BSMV (9, 13), fueron observadas en todos los preparados realizados con material de las diferentes purificaciones (figura 2).
El procedimiento implementado para purificar parcialmente al BSMV-B permitió alcanzar los resultados esperados. Aunque no se determinó el rendimiento viral a partir del tejido vegetal, los resultados de las pruebas de infectividad evidenciaron que después del proceso de precipitación con polietilenglicol (PEG) la cantidad de partículas virales que permanecieron en el sobrenadante descartado fueron escasas. Sin embargo, después de resuspender el sedimento obtenido y centrifugar, aún persistía en el nuevo sedimento que se descarta cierta cantidad de viriones (cuadro 1). Probablemente esto ocurre por restos de PEG que quedan en el sedimento resuspendido y que hace que precipiten nuevamente algunas partículas al centrifugar. Esto no afectó considerablemente el rendimiento final, ya que en la suspensión parcialmente purificada se observó una alta concentración de partículas virales, lo cual fue evidenciado a través del microscopio electrónico (figura 2) y las pruebas de infectividad (cuadro 1).
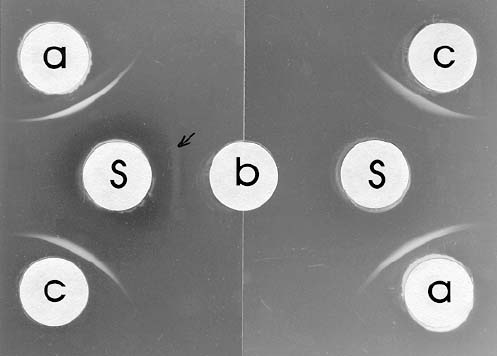
Figura 2. Partículas del BSMV-B observadas al microscopio electrónico a partir de una suspensión parcialmente purificada diluida 1:3. La barra representa 100 nm.
Cuadro 1. Pruebas de infectividad sobre Phaseolus lunatus cv Henderson Bush de algunas fracciones de la purificación parcial.1
| Fracción | Reacción2 |
| Sobrenadante descartado después de la precipitación con polietilenglicol | + |
| Ultimo sedimento descartado | ++ |
| Suspensión parcialmente purificada | +++ |
1Evaluación realizada a los cuatro días después de la inoculación mecánica. Fueron inoculadas tres plantas con cada fracción.
2+ = 1 a 2 lesiones locales necróticas(LLN)/hoja; ++ = 3 a 8 LLN/hoja. +++ = más de 9 LLN/hoja.
Estos resultados coinciden con lo citado por Christie et al.(5), quienes lograron aumentar el número de partículas observables en el microscopio electrónico mediante un protocolo sencillo de concentración para diferentes virus. La metodología desarrollada en este estudio logró ese objetivo, aparentemente en mayor cuantía, lo cual es de gran interés para la obtención de antisuero. Por otra parte, es importante destacar que durante el proceso de purificación las partículas virales no perdieron su infectividad, ya que las fracciones evaluadas indujeron síntomas característicos de la enfermedad sobre P. lunatus cv Henderson Bush (lesiones locales necróticas) y P. vulgaris cv Tacarigua (mosaico).
El título del suero normal fue cero, lo que evidencia que el conejo no presentaba anticuerpos contra un patógeno homólogo. En pruebas de doble difusión en agar se observó una banda de precipitación muy evidente entre el antígeno viral (BSMV-B) y su antisuero. Sin embargo, también se observó una banda muy tenue con proteína de planta sana (figura 3), lo que demuestra que la purificación del BSMV-B no fue total, quedando algún remanente de proteína vegetal. Este resultado era de esperarse, ya que se utilizó un protocolo sencillo de «purificación». Otros investigadores (1, 11, 14, 15) también han obtenido resultados similares con otros virus, recurriendo a la absorción del antisuero para eliminar los anticuerpos no específicos contra el virus.
El título del antisuero para el primer y segundo sangrado fue de 1:1024. Es decir, que tres inyecciones eran suficientes para obtener un suero con el mismo título. Esto es debido en gran parte a que el BSMV es altamente inmunogénico (13) y la concentración de partículas en cada inyección era alta (1 mL de suspensión viral parcialmente purificada). Como el antisuero obtenido no fue completamente específico, fue necesario absorberlo o saturarlo con proteína de planta sana. Con una dilución de 1:4 de la proteína de planta sana obtenida (utilizando el antisuero como diluente) se logró precipitar todos los anticuerpos no específicos, observándose únicamente la formación de banda de precipitación correspondiente al antígeno viral y sus anticuerpos, sin reacción simultánea con la proteína vegetal (figura 3). Después de la absorción del antisuero el título fue de 1:512.
La reacción del antisuero absorbido sin diluir fue positiva (banda de precipitación bien visible), tanto con las suspensiones virales parcialmente purificadas, como con savia de plantas infectadas. Sin embargo, en este último caso las bandas fueron ligeramente más tenues, debido a que la concentración viral era considerablemente menor con respecto a las suspensiones parcialmente purificadas (figura 3).
El antisuero absorbido permitió detectar fácilmente al BSMV-B en muestras procedentes del campo (plantas jóvenes y viejas) mediante pruebas serológicas de doble difusión en agar. Este tipo de prueba permite detectar a este virus con bastante precisión, debido a que no está relacionado serológicamente con otros miembros del grupo sobemovirus, ni con otros virus pertenecientes a otros grupos. No obstante, todas las razas del BSMV están relacionadas serológicamente entre sí (13). Por otra parte, sería interesante probar el antisuero obtenido a través de otras técnicas serológicas más sensibles, como ELISA, ya que el título del mismo es relativamente alto y se utilizaría menos antisuero.

Figura 3. Prueba de doble difusión en agar utilizando antisuero (S) contra el BSMV-B: sin absorber (izquierda) y absorbido (derecha). Los huecos periféricos contenían virus parcialmente purificado (a) y savia de plantas de caraota: sanas (b) e infectadas con BSMB-B (c).
Algunos investigadores (4, 6, 7) han obtenido antisueros contra el BSMV con títulos de 1:512, determinados mediante pruebas de doble difusión en agar. Estos sueros han sido obtenidos mediante metodologías que hacen uso de equipos especializados de alto costo, las cuales han tenido como base la precipitación de las proteínas mediante el uso de ultracentrífugas. Sin embargo, en esta investigación, con una metodología sencilla y sobre la base de la precipitación de las proteínas con polietilenglicol, se ha obtenido un antisuero con igual título, lo cual es de gran interés en nuestras condiciones. Por otra parte, sería importante evaluar este protocolo para otros virus de características similares y estudiar la posibilidad de obtener antisueros.
En conclusión, los resultados obtenidos en este trabajo coinciden con lo logrado por Alagares et al. (1), Ochoa y Trujillo (11) y Trujillo et al. (14); es decir, se pueden obtener antisueros contra algunos virus de plantas mediante técnicas simples y en laboratorios modestos. En tal sentido, la obtención de antisuero contra el BSMV-B mediante una metodología sencilla puede ser de gran ayuda en el diagnóstico y estudios sobre la epidemiología de este virus, particularmente en laboratorios o países que confronten problemas económicos para la obtención de materiales y equipos.
Agradecimientos
Los autores desean expresar su gratitud al Dr. E. Debrot, Lic. M. Alfaro y Tecn. F. Centeno (Fonaiap-Ceniap, Laboratorio de Virología Vegetal, Maracay) por su valiosa ayuda en la parte de microscopía electrónica. Igualmente, a la profesora L. Farińas (UCV, Fac. Agronomía, Laboratorio de Enzimología, Maracay), por permitir el uso de algunos equipos y materiales.
Literatura citada
1. Alagares, M., G. E. Trujillo, M. J. Garrido y C. M. de Jiménez. 1988. Algunos aspectos relacionados con el virus del mosaico severo del frijol (Vigna unguiculata (L.) Walp). Fitopatol. Venez. 1 : 25-26 (Resumen).
2. Ball, E. M. 1990. Agar double diffusion, plates (Ouchterlony): Viruses. p. 111-120. In: R. Hampton, E. Ball, and S. De Boer (Eds.). Serological methods for detection and identification of viral and bacterial plant pathogens. APS Press, Minnesota, EE.UU.
3. Ball, E. M., R. O. Hampton, S. H. De Boer, and N. W. Schaad. 1990. Polyclonal antibodies. p. 33-54. In: R. Hampton, E. Ball, and S. De Boer (Eds.). Serological methods for detection and identification of viral and bacterial plant pathogens. APS Press, Minnesota, EE.UU.
4. Cupertino, F. P, M. T. Lin, E. W. Kitajima, and C. L. Costa. 1982. Occurrence of southern bean mosaic virus in Central Brazil. Plant Disease 66 : 742-743.
5. Christie, S. R., D. E. Purcifull, W. Crawford, and N. A. Hamed. 1987. Electron microscopy of negatively stained clarified viral concentrates obtained from small tissue samples with appendices on negative staining techniques. IFAS, University of Florida, Gainesville. Bulletin 872. 45 p.
6. Grogan, R.G. and K. A. Kimble. 1964. The relationship of severe bean mosaic virus from México to southern bean mosaic virus and its related strain in cowpea. Phytopathology 54 : 74-78.
7. Lamptey, P. and R. Hamilton. 1974. A new cowpea strain of southern bean mosaic virus from Ghana. Phytopathology 64 : 1100-1104.
8. Mora, O., G. E. Trujillo, O. Borges y R. de Uzcátegui. 1988. Mosaico sureńo de la caraota (southern bean mosaic virus) afectando a Phaseolus vulgaris en Venezuela. Fitopatol. Venez. 1 : 29 (Resumen).
9. Morales, J. F. and R. Gámez. 1989. Beetletransmitted viruses. p. 363-377. In: H. F. Schwartz and M. A. Pastor-Corrales (Eds.). Bean production problems in the tropics. 2nd. ed. CIAT. Cali, Colombia.
10. Noordam, D. 1973. Identification of plant viruses. Methods and experiments. Center for Agricultural Publishing and Documentation. Wageningen, Holanda.
11. Ochoa, F. y G. E. Trujillo. 1994. Metodología sencilla para la obtención de antisuero al virus de la tristeza de los cítricos. Fitopatol. Venez. 7 : 2-5.
12. Patińo, Y. y M. J. Garrido. 1996. Identificación y detección del virus del mosaico sureńo de la caraota en varias localidades del estado Aragua, Venezuela. Fitopatol. Venez. 9 : 16-17.
13. Tremaine, J. H. and R. I. Hamilton. 1983. Southern bean mosaic virus. Descriptions of plant viruses No. 274. CMI/AAB. Kew, Surrey, England. 6 p.
14. Trujillo, G. E., M. J. Garrido y M. Alagares. 1989. Un modo fácil de obtener antisueros a diferentes virus vegetales. p. 21-22. En: Memoria II Jornadas de Investigación de la Facultad de Agronomía, UCV. Maracay, Venezuela.
15. Van Regenmortel, M. H. V. 1982. Serology and inmunochemistry of plant viruses. New York, Academic Press.